 |
 |
 |
 |
 |
|
| Koi | Penguin | Whirligig Beetle |
Wasp Model |
Beetle Mimic |
Evolution is the change in frequencies of the various alleles for some gene in a given population through time. These changes in allelic frequencies are brought about by the effects of natural selection and may lead, eventually, to speciation. Thus, we need to examine, What is a species? and How does one species become two (or more) species?
In 1942, Ernst Mayr defined a species as a group of actually or potentially interbreeding populations that are reproductively isolated from other such groups.
The term isolating mechanisms refers to any means by which the many, diverse species remain distinct. These may be morphological (the bodies dont fit together), behavioral (day vs. night, spring vs. fall, courtship rituals), and/or geographical (separated by mountain, desert, lake, river, ocean, or other barrier) barriers. There are several different types of species, based on the mechanism(s) which keep those species isolated from other, similar species.
|
|
|
|
|
|
| Koi | Penguin | Whirligig Beetle |
Wasp Model |
Beetle Mimic |
Within a species there is genetic variation due to mutations and crossing over in meiosis (as recognized by Darwin and Wallace). These variations may be grouped into several categories.
 |
 |
| Male Downy Woodpecker |
Female Downy Woodpecker |
Because of this variation, some individuals area better suited to cope with their environments than others, or in other words, the environment provides selective pressure on the species/population. Two false assumptions students typically make are worth noting:
Rather, as stated by Darwin and Wallace, random mutations just happen, and some make the organism better or worse able to get along, given existing environmental conditions.
Natural selection is the differential reproductive success of individual organisms (given existing environmental conditions), or in other words, the ability of individuals to
The relationship between sickle-cell anemia and malaria is an interesting example of natural selection. Sickle-cell is caused by a mutation in one nucleotide in the sequence which codes for hemoglobin, resulting in one different amino acid in the sequence, and thereby causing the persons RBCs to sickle under stress (which leads to other problems). Sickle-cell is a recessive allele, but unlike other recessive alleles, it does not code for I dont know how to make this, but rather Im going to make this, but make it wrong. Thus, an individual who is SS will make all normal hemoglobin, a person who is Ss has two sets of instructions and will make both normal and sickle-cell hemoglobin, and a person who is ss will make only abnormal, sickle-cell hemoglobin. The rather pointed, jagged, sickled RBCs of a person with sickle-cell tend to get caught in capillaries and trigger blood clots. Thus, many people who are ss (sickle-cell disease) die from complications of the sickle-cell (natural selection against homozygous recessive individuals).
However, malaria is also present in the part of the world where the sickle-cell allele arose, and as it turns out, sickle-cell confers resistance against the malaria parasite. When a malaria parasite invades a normal RBC, it then goes on to reproduce and invade other RBCs, but when a malarial parasite invades a RBC containing abnormal, sickle-cell hemoglobin, the cell is put under so much stress by the presence of the parasite, that the cell sickles (crinkles up), killing the parasite in the process. Thus, normal SS people often die due to causes associated with the malaria (natural selection against the homozygous dominant). Because Ss (sickle-cell trait) individuals have some normal hemoglobin, it takes a higher level of stress on their systems to cause their RBCs to sickle. Also, they are somewhat resistant to malaria (RBCs containing abnormal hemoglobin will sickle if attacked, but RBCs with regular hemoglobin wont). Thus, most of the people in those populations are heterozygotes, and in that environment, end up being more fit than either of the homozygotes.
According to various sources (including the Merck Manual), in some areas of Africa 64% of the people are SS, 32% are Ss, and 4% are ss. From this, it is possible to calculate that the selective pressure against ss is 4 × the selective pressure against SS (4 × more people die from sickle-cell than from malaria). However, among African Americans, 90.25% of the population is SS, 9.50% is Ss, and 0.25% is ss. From this, it is possible to calculate that the selective pressure against ss is 19 × the selective pressure against SS (19 × more people die from sickle-cell than from malaria). Comparatively more people here die from sickle-cell vs. from malaria than in Africa (fewer people here die from malaria).
In discussing natural selection, a textbook we were using several years ago says that, Those individuals that contribute the most to the gene pool are said to be the most fit, and those that contribute little or nothing to the gene pool are said to be the least fit. The fitness of an individual is measured by its reproducing offspring. . . In general, while dealing with wild organisms this seems to make sense, but an interesting rhetorical question or paradox arises if this is applied to humans. Ironically, we criticize rapidly-growing poor or Third-World populations, virtually telling them that if they were as smart, as well off, or whatever, as us (read: as fit as) that they should have less offspring like we do. Here in Western Civilization, despite lower rates of diseases, more food, better medical care, etc., the reproduction rate in some countries is not even high enough to maintain current population levels, and population sizes are actually declining. Here, then, is something to consider: just who is the most fit?
Reference: Smith, Robert Leo 1992. Elements of Ecology, 3rd Ed. HarperCollins Publ., NY. p. 18.
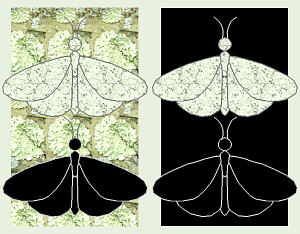 One of the best documented examples of natural selection in modern times is
the English Peppered Moth, Biston betularia. Typically, this moth is
whitish with black speckles and spots all over its wings. During the daytime,
Peppered Moths are well-camouflaged as they rest on the speckled lichens on
tree trunks. Occasionally a very few moths have a genetic mutation which
causes them to be all black, so they are said to be
melanistic.
Black moths resting on light-colored, speckled lichens are not very well
camouflaged, and so are easy prey for any moth-eating birds that happen by.
Thus, these melanistic moths never get to reproduce and pass on their genes
for black color. However, an interesting thing happened to these moths in
the 1800s. With the Industrial Revolution, many factories and homes in
British cities started burning coal, both for heat and to power all those
newly-invented machines. Coal does not burn cleanly, and creates a lot of
black soot and pollution. Since lichens are extremely sensitive to air
pollution, this caused all the lichens on city trees to die. Also, as the
soot settled out everywhere, this turned the tree trunks (and everything
else) black. This enabled the occasional black moths to be well-camouflaged
so they could live long enough to reproduce, while the normal speckled
moths were gobbled up. Studies done in the earlier 1900s showed that while
in the country, the speckled moths were still the predominant form, in the
cities, they were almost non-existant.
One of the best documented examples of natural selection in modern times is
the English Peppered Moth, Biston betularia. Typically, this moth is
whitish with black speckles and spots all over its wings. During the daytime,
Peppered Moths are well-camouflaged as they rest on the speckled lichens on
tree trunks. Occasionally a very few moths have a genetic mutation which
causes them to be all black, so they are said to be
melanistic.
Black moths resting on light-colored, speckled lichens are not very well
camouflaged, and so are easy prey for any moth-eating birds that happen by.
Thus, these melanistic moths never get to reproduce and pass on their genes
for black color. However, an interesting thing happened to these moths in
the 1800s. With the Industrial Revolution, many factories and homes in
British cities started burning coal, both for heat and to power all those
newly-invented machines. Coal does not burn cleanly, and creates a lot of
black soot and pollution. Since lichens are extremely sensitive to air
pollution, this caused all the lichens on city trees to die. Also, as the
soot settled out everywhere, this turned the tree trunks (and everything
else) black. This enabled the occasional black moths to be well-camouflaged
so they could live long enough to reproduce, while the normal speckled
moths were gobbled up. Studies done in the earlier 1900s showed that while
in the country, the speckled moths were still the predominant form, in the
cities, they were almost non-existant.
 Nearly all the moths in the cities were the black form. It was evident to
the researchers studying these moths that the black city moths were breeding
primarily with other black city moths while speckled country moths were
breeding primarily with other speckled country moths. Because of this, any
new genetic mutations in one or the other of those populations would only be
passed on within that population and not throughout the whole moth population.
Additionally, because the city and country environments were different, there
were different selective pressures on city vs. country moths that could
potentially drive the evolution of these two populations of moths in
different directions. The researchers pointed out that if this were to
continue for a long enough time, the city and country moths could become so
genetically different that they could no longer interbreed with each other,
and thus would be considered distinct species. In this case, what actually
happened is that the people of England decided they didnt like breathing
and living in all that coal pollution, thus found ways to clean things up.
As the air became cleaner, lichens started growing on city trees again, thus
the direction of the selective pressure (birds) was once again in favor of the
speckled moths. By now, English cities, as well as countrysides, all have
speckled moths, and all those moths are interbreeding at random, thus they
were not separated for long enough to develop into separate species.
Nearly all the moths in the cities were the black form. It was evident to
the researchers studying these moths that the black city moths were breeding
primarily with other black city moths while speckled country moths were
breeding primarily with other speckled country moths. Because of this, any
new genetic mutations in one or the other of those populations would only be
passed on within that population and not throughout the whole moth population.
Additionally, because the city and country environments were different, there
were different selective pressures on city vs. country moths that could
potentially drive the evolution of these two populations of moths in
different directions. The researchers pointed out that if this were to
continue for a long enough time, the city and country moths could become so
genetically different that they could no longer interbreed with each other,
and thus would be considered distinct species. In this case, what actually
happened is that the people of England decided they didnt like breathing
and living in all that coal pollution, thus found ways to clean things up.
As the air became cleaner, lichens started growing on city trees again, thus
the direction of the selective pressure (birds) was once again in favor of the
speckled moths. By now, English cities, as well as countrysides, all have
speckled moths, and all those moths are interbreeding at random, thus they
were not separated for long enough to develop into separate species.
Dr. H. B. D. Kettlewell, the main researcher who worked on Peppered Moths discovered the following:
Reference: Kettlewell, H. B. D. 1961. The phenomenon of industrial melanism in Lepidoptera. Ann. Rev. of Entomol. 6: 245 - 262.
However, an article in the 24 May 1999 issue of The Scientist 13(11) presents and discusses data which may refute this long-held idea.
An organisms alleles are subjected to natural selection. Some of the alleles code for certain body features or behaviors which are better suited to the environment than others. Organisms with those alleles are more likely to reproduce and pass on those alleles. Other alleles are not as well suited, and an organism with that genotype dies or is not able to reproduce, so the alleles never get passed on. Over time, this leads to speciation, the division of one species into two or more species.
Natural selection may, then, lead to cladogenesis, which is the splitting of one species into two or more species, in other words, speciation. There are several theories on exactly how this happens. Some biologists feel that the most common means is gradual speciation, in which allopatric or geographically isolated populations of an organism, each subject to unique selective pressures (climate, etc.), gradually become so genetically different that they can no longer interbreed, even if the geographical barrier is subsequently removed. This long-held belief is based on the works of Darwin, Wallace, and others who said that species are always slowly evolving. In contrast, abrupt speciation as part of punctated equilibrium is an instantaneous speciation (taking thousands of years rather than tens of thousands or millions), followed by long periods of species stability (with relatively little change to the gene pool). This fairly-recent and somewhat controversial theory, proposed by paleontologists to explain holes in the fossil record, postulates that new species may arise spontaneously (for example, via polyploidy), and then are fairly stable.
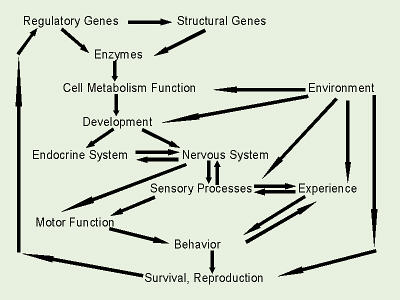 One important thing to note here is that populations, not
individuals, evolve: individuals can have genetic mutations, but unless
those mutations are passed along to offspring and thus into the gene
pool of the population, no real change has occurred. An individual who
has a mutation but does not pass it on (for whatever reason) has no long
term effect on the genes present in the population as a whole. Another
important point is that natural selection operates on the genetics of
a species, including whatever genes affect instinctive behavior patterns as
well as those affecting the looks of the individuals. Thus, genetics, looks,
behavior, and external environmental factors are all interwoven.
One important thing to note here is that populations, not
individuals, evolve: individuals can have genetic mutations, but unless
those mutations are passed along to offspring and thus into the gene
pool of the population, no real change has occurred. An individual who
has a mutation but does not pass it on (for whatever reason) has no long
term effect on the genes present in the population as a whole. Another
important point is that natural selection operates on the genetics of
a species, including whatever genes affect instinctive behavior patterns as
well as those affecting the looks of the individuals. Thus, genetics, looks,
behavior, and external environmental factors are all interwoven.
 Sometimes,
there are tradeoffs between various genes and selective pressures. For
example, the bright red color of a male Cardinal might be very attractive to
a female, such that the brighter a male is, the greater his chances of mating,
but that same bright red color, especially against a snowy background, makes
him much more visible to predators.
Sometimes,
there are tradeoffs between various genes and selective pressures. For
example, the bright red color of a male Cardinal might be very attractive to
a female, such that the brighter a male is, the greater his chances of mating,
but that same bright red color, especially against a snowy background, makes
him much more visible to predators.
Species of organisms may spread out into nearby, available habitats (and if the selective pressure is different there, may eventually give rise to another species). Adaptive radiation is the process whereby a species spreads into and makes use of a new environment. In order for this to occur, several conditions must be met:
Probably one of the most famous and best studied examples of adaptive radiation is Darwins finches. It is thought that these 14 species of birds on the Galapagos Islands arose from a few birds of one species which originally came from the Ecuadorian mainland and eventually adapted to a variety of diets and habitats. Another similar example involves a number of species of fruit flies on the Hawaiian Islands.