You may type in your own answers, then check to see if
you were right. If youre totally stumped, you can tell the computer to
show you the answer to a particular question.
Monohybrid Cross:
In humans, brown eyes (B) are dominant over blue (b)*. A brown-eyed man
marries a blue-eyed woman and they have three children, two of whom are
brown-eyed and one of whom is blue-eyed. Draw the Punnett square that
illustrates this marriage. What is the mans genotype? What are the
genotypes of the children?
(* Actually, the situation is complicated by
the fact that there is more than one gene involved in eye color, but for this
example, well consider only this one gene.)
Testcross:
In dogs, there is an hereditary deafness caused by a recessive gene, d. A
kennel owner has a male dog that she wants to use for breeding purposes if
possible. The dog can hear, so the owner knows his genotype is either DD or
Dd. If the dogs genotype is Dd, the owner does not wish to use him for
breeding so that the deafness gene will not be passed on. This can be tested
by breeding the dog to a deaf female (dd). Draw the Punnett squares to
illustrate these two possible crosses. In each case, what percentage/how
many of the offspring would be expected to be hearing? deaf? How could you
tell the genotype of this male dog? Also, using Punnett square(s), show how
two hearing dogs could produce deaf offspring.
Incomplete Dominance:
Note: at
least one textbook Ive seen also uses this as an example of pleiotropy (one
gene multiple effects), though to my mind, the malaria part of this is not
a direct effect of the gene.
(For many genes, such as the two mentioned above, the dominant allele codes
for the presence of some characteristic (like, B codes for make brown
pigment in someones eyes), and the recessive allele codes for something
along the lines of, I dont know how to make that, (like b codes for the
absence of brown pigment in someones eyes, so by default, the eyes
turn out blue). If someone is a heterozygote (Bb), that person has one set
of instructions for make brown and one set of instructions for, I dont
know how to make brown, with the result that the person ends up with brown
eyes. There are, however, some genes where both alleles code for
something. One classic example is that in many flowering plants such as
roses, snapdragons, and hibiscus, there is a gene for flower color with two
alleles: red and white. However, in that case, white is not merely the
absence of red, but that allele actually codes for, make white pigment.
Thus the flowers on a plant that is heterozygous have two sets of
instructions: make red, and make white, with the result that the
flowers turn out mid-way in between; theyre pink.)
In humans, there is a gene that controls formation of hemoglobin, the protein
in the red blood cells which carries oxygen to the body tissue.
The normal allele of this gene codes for normal hemoglobin. However,
there is another allele for this gene that has one different nitrogenous
base in its DNA sequence, and thus, one codon in the middle of the gene
codes for a different amino acid in an important place in the hemoglobin
molecule. A red blood cell (RBC) that contains this altered hemoglobin will,
under stress, crinkle up into a shape that reminded someone of the shape of
an old-fashioned sickle. While the letters S ands are often used to
represent these alleles, since both of them code for make hemoglobin, in
reality, neither is dominant over the other. Someone who is SS makes all
normal hemoglobin, someone who is ss makes all abnormal hemoglobin (and we
say that person has sickle-cell anemia), and someone who is Ss essentially
has two sets of instructions, and so, makes some of each kind of hemoglobin
(often referred to as sickle-cell trait).
Because the RBCs of a person who is ss contain all abnormal hemoglobin, they
will sickle very easily, with very little stress required to provoke that
reaction. All those sickled cells tend to get stuck as they try to go through
capillaries, and cause things like strokes, heart attacks, pulmonary embolisms,
etc. that lead to death. Because only some of the RBCs of a person who is Ss
contain abnormal hemoglobin, that person usually only has trouble with a lot
of cells sickling if theyre under a lot of stress trying to meet a
higher-than-normal oxygen demand, and so the chances of a
person dying from sickle-cell trait are much lower than for full-blown
sickle-cell anemia.
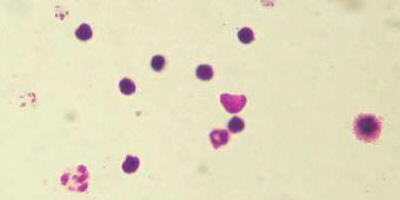
A photo, taken by Dr. Fankhauser, of a prepared slide of blood
cells infected with Plasmodium vivax
Malaria is a parasitic disease thats prevalent in tropical areas. When a
mosquito thats carrying the parasites bites someone, the parasites enter
the persons bloodstream, and invades and lives in the persons RBCs. However,
if a person has sickle-cell anemia (ss), the presence of a parasite in a RBC
is so stressful, it causes the RBC to sickle (crinkle up), and when that happens,
that kills the parasite before it can multiply and spread to other RBCs. Thus,
coincidentally, a person who is ss is also immune to malaria. If a person
is Ss and a malaria parasite tries to invade a RBC with abnormal hemoglobin,
again, the RBC will sickle, killing the parasite before it has a chance to
reproduce. If a parasite invades a RBC with normal hemoglobin, it
will be able to live and multiply, but if its offspring invade other RBCs with
abnormal hemoglobin, they, too, will be killed. Thus, a person who is Ss is
resistant (though not totally immune) to malaria. If a person is SS and has
all normal hemoglobin, the malaria parasites do just fine, invading RBCs,
growing and multiplying, and invading more RBCs. Thus, an SS person usually
dies, eventually, from causes tied to the malaria.
A man and woman living in a tropical area where malaria
is prevalent and health care is not accessible have seven children. The
genotypes of these children are ss, Ss, SS, ss, Ss, Ss, and SS.
Dihybrid Cross:
In humans, there is a gene that controls formation (or lack
thereof) of
muscles in the tongue that allow people with those muscles to roll their
tongues, while people who lack those muscles cannot roll their tongues. The
ability to roll ones tongue is dominant over non-rolling. The ability to
taste certain substances is also genetically controlled. For example, there
is a substance called phenylthiocarbamate (PTC for short), which some people
can taste (the dominant trait), while others cannot (the recessive trait).
The biological supply companies actually sell a special kind of tissue paper
impregnated with PTC so students studying genetics can try tasting it to see
if they are tasters or non-tasters. To people who are tasters, the paper
tastes very bitter, but to non-tasters, it just tastes like paper. Lets let
R represent tongue-rolling, r represent a non-roller, T
represent ability to taste PTC, and t represent non-tasting.
Suppose a woman who is both a homozygous tongue-roller and a non-PTC-taster
marries a man who is a heterozygous tongue-roller and is a PTC taster, and
they have
three children: a homozygous tongue-roller who is also a PTC taster, a heterozygous
tongue-roller who is also a taster, and a heterozygous tongue-roller who is a
non-taster. If these parents would have a bunch more children so that they
had 12 in all, how many of those 12 would you expect to be non-tasters who
are homozygous for tongue-rolling? If the first child (the homozygous
tongue-roller who is also a PTC taster) marries someone who is heterozygous
for both traits, draw the Punnett square that predicts what their children
will be.
If the man is both Rr and Tt (How do we know that?), he would be
RrTt and so could produce gametes with either R or r and either T or t
(one allele for each gene). There are two choices for
the first trait (R or r). No matter which of those go into a given sperm,
there are still two choices for the second trait (T or t). Therefore, a total
of 2 × 2 = 4 possible types of gametes (sperm) may be produced.
Multiple Alleles and Codominance:
Some genes have more than two alleles. One of the best-known
examples is
the gene that is referred to as the ABO Blood Group, which actually has
quite a number of alleles. However, we will discuss/consider only the three
most-common of these. This gene codes for the structure of a certain antigen
on the surface of our RBCs. The three alleles we will work with are symbolized
by IA, IB, and i. However, keep in mind that a person
can only have two alleles, two copies of a gene. Thus, the possible genotypes
are IAIA, IAi, IBIB,
IBi, IAIB, or ii. (Sometimes, you will see
these simplified as AA, AO, BB, BO, AB, and OO, but that does make it harder
to remember that these are all alleles for the same gene.)
The allele, IA, codes for, make type A antigen, the allele
IB codes for, make type B antigen, and (to simplify things
somewhat) the i allele codes for, I dont know how to make either A or B.
Thus, both IAIA and IAi individuals receive
instructions to make type A antigen, and both IBIB and
IBi individuals receive instructions to make type B antigen.
Individuals who are IAIB receive two sets of instructions:
make type A and make type B, so they have both the A and B forms
of that antigen on the surface of their RBCs.
People who are ii dont have any instructions to make either A or B, so by
default they make what we refer to as type O antigens. Since both IA
and IB code for make something whereas i codes for, I dont know
how, therefore, both IA and IB are dominant over i.
However, since both IA and IA code for make something,
neither of them is dominant over the other. Thus we say that IA
and IB are codominant over i.
Suppose a person with type A blood and a person with type B blood get married.
What are the possible genotypes their children could have?
There is another gene that codes for another, different antigen that also
occurs on the surface of our RBCs, and technically, that gene also has multiple
alleles. However, most people either have or do not have one particular allele
called the d allele. This gene codes for an antigen that is called Rh factor
because it was first discovered in Rhesus monkeys. People who have
instructions to make d antigen are referred to as Rh+ (the allele
is often symbolized by the letter R), while those who have I dont know
how to make d antigen instructions are called Rh (the allele can be
symbolized by r). Since this is a totally separate gene than the ABO blood
group, if youre doing a genetic cross that involves both ABO and Rh,
that would be a dihybrid cross.
Ms. Johnston, Ms. Johnson, and Ms. Johnstone all entered the same hospital and
gave birth to baby girls on the same day, and all three babies were taken to
the nursery to receive care, there. Someone later claimed that the hospital
mixed up the babies. As a hospital administrator, it is your job to make
sure that each pair of parents has the correct baby, so you order blood typing
to be done on all the parents and all the babies. Here are the results:
Trihybrid Cross:
In Guinea pigs, black hair (B) is dominant over white (b),
rough coat texture (R) is dominant over smooth (r), and short hair (S) is
dominant over long hair (s). Assuming these genes are on separate chromosomes,
draw the Punnett square for a cross between a homozygous black, rough,
short-haired Guinea pig and a white, smooth, long-haired one. What would the
phenotype(s) of the offspring be? If two of the F1 offspring
were crossed, draw the Punnett square for this cross. Hint: first make a
list of the possible gametes, making sure each has exactly one copy
of each of the genes (one allele for each gene). What would
the genotype and phenotype ratios be for the F2 generation?
Polygenic Trait:
Some traits, some phenotypes, are controlled by more than one
gene. It was
mentioned in the monohybrid cross, above, that technically, human eye color
is controlled by at least two genes, one which codes for brown vs. blue and
another which codes for green vs. blue. In the epistasis crosses, below,
you will see other examples of polygenic traits. Human skin color is also
a classic example of a polygenic trait. It is known that at least three or
four genes control skin color, and for each of those genes, dark pigment has
incomplete dominance over light (so a heterozygote would be intermediate see
above). Because we just did a trihybrid cross, lets assume three genes
here (for simplicity), and to avoid confusion among them, lets arbitrarily
call them genes A, B, and C. Then, someone who is
AABBCC
would have very dark skin color and someone who is
aabbcc
would have very light skin color. If they would get married and have children,
their children would all be
AaBbCc.
If two AaBbCc
people would get married and have children, the Punnett square would look
like this:
ABC
ABc
AbC
Abc
aBC
aBc
abC
abc
ABC
AABBCC
AABBCc
AABbCC
AABbCc
AaBBCC
AaBBCc
AaBbCC
AaBbCc
ABc
AABBCc
AABBcc
AABbCc
AABbcc
AaBBCc
AaBBcc
AaBbCc
AaBbcc
AbC
AABbCC
AABbCc
AAbbCC
AAbbCc
AaBbCC
AaBbCc
AabbCC
AabbCc
Abc
AABbCc
AABbcc
AAbbCc
AAbbcc
AaBbCc
AaBbcc
AabbCc
Aabbcc
aBC
AaBBCC
AaBBCc
AaBbCC
AaBbCc
aaBBCC
aaBBCc
aaBbCC
aaBbCc
aBc
AaBBCc
AaBBcc
AaBbCc
AaBbcc
aaBBCc
aaBBcc
aaBbCc
aaBbcc
abC
AaBbCC
AaBbCc
AabbCC
AabbCc
aaBbCC
aaBbCc
aabbCC
aabbCc
abc
AaBbCc
AaBbcc
AabbCc
Aabbcc
aaBbCc
aaBbcc
aabbCc
aabbcc
Linked Genes:
wild-type female
wild-type male
black, vestigial female
black, vestigial male
Normal fruit flies have grayish-yellow bodies, red eyes, and wings that are
long-enough to be able to fly. Some mutant fruit flies have black bodies rather
than grayish-yellow, some have stumpy, vestigial wings that are too short and
wrinkled to be able to fly, and some have a brighter, orangish-red eye color
that is called cinnabar. By breeding flies, fruit fly researchers were able
to determine that all three of these mutations were recessive and were on the
autosomes.
Fruit fly researchers use a different type of symbolism to represent their
genetic crosses. They use a plus sign (+) to indicate anything that is the
wild type and a letter (or sometimes two) to represent a mutant allele
(capital if the mutation is a dominant allele, lower case if its recessive).
Thus a fruit fly with a homozygous grayish-yellow body would be labeled as
++ for body color while a black-bodied fly would be bb and a heterozygous
fly would be +b. The genotype for normal wings would also be symbolized as
++, while the allele for vestigial wings is vg so a fly that is
homozygous recessive for that gene would be vgvg and a heterozygote would
be notated as +vg. Similarly, normal red eyes would be ++, cinnabar eyes
would be cncn, and a heterozygote would be +cn.
To simplify things, for now, lets consider just the b and vg genes. Suppose
we had some flies that were genotype +b+vg (grayish-yellow body and long
wings, but carriers for both the black and vestigal alleles). Lets do a
testcross (= a cross with bbvgvg, remember?) with these heterozygous flies.
If these genes were on different chromosomes, the Punnett
square for this cross would look like this:
++
+vg
b+
bvg
bvg
+b+vg
+bvgvg
bb+vg
bbvgvg
so out of the offspring, we would expect to get ¼ of each of
the four types.
If the genes were linked on the same chromosome and one of
the flys
chromosomes contained the alleles for yellowish body and normal wings while
the other chromosome contained the alleles for black body and crumpled,
vestigial wings, the Punnett square for this cross would look like
this:
++
bvg
bvg
+b+vg
bbvgvg
so out of the offspring, we would expect to get ½ of each of
the two types.
Sex-Linked Genes:
In humans, the genes for colorblindness and hemophilia are
both located on the X chromosome with no corresponding gene on the Y.
Typically an X chromosome with a normal allele is notated as a plain X
while an X chromosome carrying the mutant allele is notated as X′
or with an appropriate letter, such as Xc for colorblindness
or Xh for hemophilia (thus, while its not typically used,
you could use XC or XH to represent the
normal allele, especially if youre dealing with both, simultaneously). Note
that besides being sex-linked genes, hemophilia and colorblindness are also
linked genes and a man who has both would be XchY.
These
are both recessive alleles. If a man and a woman, both with normal vision,
marry and have a colorblind son, draw the Punnett square that illustrates this.
If the man dies and the woman remarries to a colorblind man, draw a Punnett
square showing the type(s) of children could be expected from her second
marriage. How many/what percentage of each could be expected?
Sex (Its a Phenotype, Not a Chromosome!):
In humans, theres another X-linked gene that codes for
androgen (testosterone)
receptors in our cells. The dominant allele (XA) codes for make
functioning receptors that can correctly receive and bind onto testosterone,
but there is a recessive allele (Xa) that either codes for I dont
know how to make testosterone receptors or else make broken receptors that
cant receive and bind onto testosterone. In humans, there is also a Y-linked
gene that codes for make testes, and when present, they, in turn, make
testosterone, and the testosterone, in turn, goes to other cells in the body,
and when received by the receptors, triggers other events within those cells.
During embryonic development, one such event is growth and development of
male genitalia.
Consider a person who is genotype XAY. Because this person has
a Y chromosome including a normally-functioning gene to make testes, at the
appropriate time during embryonic development, testes will form and will
start to secrete testosterone. Because this person also has the
correctly-functioning allele for the androgen (testosterone) receptor gene,
those receptors will form and will begin functioning. As they receive the
testosterone made by the testes, this will stimulate development of male
genitalia, and (assuming all other genes are working normally),
this baby will be a boy.
Consider a person who is genotype XAXA. Because this
person does not have a Y chromosome, there is no gene to provide instructions
to make testes, therefore no big prenatal surge of testosterone, therefore
no stimulus to make male genitalia (in spite of properly-functioning
testosterone receptors), so by default, female genitalia develop, and
(assuming all other genes are working normally), this baby will be
a girl.
If these two people would get married, the Punnett square
for their children would look pretty much like the ones youve just done:
XA
Y
XA
XAXA
XAY
XA
XAXA
XAY
½ of the children will be females, and ½ will be males
Consider a person who is genotype XAXa. Because this
person does not have a Y chromosome, there is no gene to provide instructions
to make testes, therefore no big prenatal surge of testosterone, therefore
no stimulus to make male genitalia (in spite of properly-functioning
testosterone receptors in many/most cells), so by default, female genitalia
develop, and (assuming all other genes are working normally), this baby
will also be a girl. However, she is a carrier for the non-functioning
androgen-receptor gene. Because of inactivation of X chromosomes to form
Barr bodies, some of the cells in her body will not be able to receive
testosterone messages, which might show up in things like having
somewhat less pubic and/or armpit hair.
Sex-Influenced Traits:
Baldness in humans is a dominant, sex-influenced trait. This gene is on the
autosomes, not the sex chromosomes, but how it is expressed is influenced
by the persons sex (due to hormones present, etc.). A man who is BB or Bb
will be bald and will be non-bald only if he is bb. A woman will only be
bald if she is BB and non-bald if she is Bb or bb (its almost like B is
dominant in males and b is dominant in females). Actually, because of the
influence of other sex-related factors, most women who are BB never become
totally bald like men do, but rather, their hair may become thin or sparse.
If two parents are heterozygous for baldness (note: that means he would be
bald, but she would not.), what are the chances of their children being
bald? Use a Punnett square to illustrate this.
Note: because the sex of a person does make a difference in how the
gene is expressed, you need to set this up as a dihybrid cross to account
for the sex of the children.
A non-bald man marries a non-bald woman. They have a son and
a daughter. If the son becomes bald, what are the chances that his sister
will, too? Use a Punnett square to show this cross.
Epistasis:
In sweet peas, purple flower color (P) is dominant over white (p), but there
is also a control gene such that if the plant has a C, the purple has
permission to express itself. If the plant is cc, the purple does not
have permission to express itself and the flower will be white anyway. If
a plant with homozygous purple, controlled flowers is crossed with a plant
with white, non-controlled flowers, diagram the Punnett square for the
F1 and F2 generations and calculate the genotype and
phenotype ratios.
Because you must keep track of both the purple/white and the control genes,
you need to set this up as a dihybrid cross.
Epistasis can also work in the opposite direction (actually,
some textbooks give these two cases other names). In corn kernels, purple
(P) is dominant over yellow (p), but there is also an inhibiting control gene
such that if that corn kernel has a C, the purple does not have permission
to express itself, and the kernel will be yellow. If that kernel is cc,
then the purple has permission to be expressed. If the same cross as above
is done, the result would be:
genotypes:
phenotypes:
PPCC
1
PC
= yellow bec. of C
9
}
13 yellow total
PPCc
2
ppC
= yellow
3
PPcc
1
ppcc
= yellow bec. of pp
1
PpCC
2
Pcc
= purple
3
}
3 purple
PpCc
4
Ppcc
2
ppCC
1
ppCc
2
ppcc
1
There are some other, interesting possibilities. Suppose that
in corn kernels, purple (P) is dominant over some other color, say red (p).
However, suppose there is a control gene (C) which permits the color to be
present and cc doesnt permit color to be present, resulting in yellow
kernels. If the same cross as above is done, the result would be:
genotypes:
phenotypes:
PPCC
1
PC
= purple
9
}
9 purple
PPCc
2
Pcc
= yellow bec. of cc
3
}
4 yellow total
PPcc
1
ppcc
= yellow bec. of cc
1
PpCC
2
ppC
= red bec. of pp
3
}
3 red
PpCc
4
Ppcc
2
ppCC
1
ppCc
2
ppcc
1
Suppose, instead, that C inhibits or does not permit the
color to be present, while cc does allow the color to be expressed.
If the same cross as above is done, the result would be:
genotypes:
phenotypes:
PPCC
1
PC
= yellow bec. of C
9
}
12 yellow total
PPCc
2
ppC
= yellow bec. of C
3
PPcc
1
Pcc
= purple
3
}
3 purple
PpCC
2
ppcc
= red bec. of pp
1
}
1 red
PpCc
4
Ppcc
2
ppCC
1
ppCc
2
ppcc
1
ZW Sex-Linked Inheritance:
Birds and lepidopterans (butterflies and moths) have ZW sex
determination, which is so named to distinguish it from XY sex determination
(as in humans and fruit flies) because ZW is the opposite of XY. In the ZW
sex determination system, the females are heterogametic (ZW), and the males
are homogametic (ZZ).
Barred Rooster
In chickens, there is an autosomal gene called extended black, symbolized
by E. There is also a sex-linked (Z-linked) dominant gene for barred
feathers (causing the feathers of the adults to be black and white
striped/barred). Since this is a sex-linked gene, the barred allele
would be symbolized as ZB, and the non-barred allele as
Zb.
A genetic cross that is routinely done in the poultry industry is to cross
a Rhode Island Red or New Hampshire breed rooster, both of which are
EEZbZb for these two genes with a Barred Plymouth
Rock hen, which breed is EEZBW for these two genes. This
results in hybrid offspring of a variety (not a true-breeding breed) called
Black Sex-Linked. When the new Black Sex-Linked chicks hatch, they can
be sexed immediately because the females are all black while the males
have a cream-colored or whitish spot on their heads. As adults, the hens
will be solid-colored while the roosters will have barred feathers.
For purposes of this illustration, since both parents are EE, we can
ignore that gene, and look just at the sex-linked (Z-linked) barring gene.
Thus, the cross to produce Black Sex-Linked chicks can be represented by
ZBW × ZbZb.
The resulting offspring would be ZbW hens (called pullets when
young) and ZBZb roosters (called cockerels when young).
It is the ZB allele inherited by the roosters that gives them
their white head spots as chicks and also give them barred feathers as
adults.
If one each of those Black Sex-Linked hens and roosters would
manage to live to adulthood and mate with each other:
ZB
Zb
Zb
ZBZb
ZbZb
W
ZBW
ZbW
Thus, with the probability of ¼ of each of four possible types
of offspring, clearly this hybrid variety of chickens is not
true-breeding.
 In humans, brown eyes (B) are dominant over blue (b)*. A brown-eyed man
marries a blue-eyed woman and they have three children, two of whom are
brown-eyed and one of whom is blue-eyed. Draw the Punnett square that
illustrates this marriage. What is the mans genotype? What are the
genotypes of the children?
In humans, brown eyes (B) are dominant over blue (b)*. A brown-eyed man
marries a blue-eyed woman and they have three children, two of whom are
brown-eyed and one of whom is blue-eyed. Draw the Punnett square that
illustrates this marriage. What is the mans genotype? What are the
genotypes of the children?  In dogs, there is an hereditary deafness caused by a recessive gene, d. A
kennel owner has a male dog that she wants to use for breeding purposes if
possible. The dog can hear, so the owner knows his genotype is either DD or
Dd. If the dogs genotype is Dd, the owner does not wish to use him for
breeding so that the deafness gene will not be passed on. This can be tested
by breeding the dog to a deaf female (dd). Draw the Punnett squares to
illustrate these two possible crosses. In each case, what percentage/how
many of the offspring would be expected to be hearing? deaf? How could you
tell the genotype of this male dog? Also, using Punnett square(s), show how
two hearing dogs could produce deaf offspring.
In dogs, there is an hereditary deafness caused by a recessive gene, d. A
kennel owner has a male dog that she wants to use for breeding purposes if
possible. The dog can hear, so the owner knows his genotype is either DD or
Dd. If the dogs genotype is Dd, the owner does not wish to use him for
breeding so that the deafness gene will not be passed on. This can be tested
by breeding the dog to a deaf female (dd). Draw the Punnett squares to
illustrate these two possible crosses. In each case, what percentage/how
many of the offspring would be expected to be hearing? deaf? How could you
tell the genotype of this male dog? Also, using Punnett square(s), show how
two hearing dogs could produce deaf offspring.




 Baldness in humans is a dominant, sex-influenced trait. This gene is on the
autosomes, not the sex chromosomes, but how it is expressed is influenced
by the persons sex (due to hormones present, etc.). A man who is BB or Bb
will be bald and will be non-bald only if he is bb. A woman will only be
bald if she is BB and non-bald if she is Bb or bb (its almost like B is
dominant in males and b is dominant in females). Actually, because of the
influence of other sex-related factors, most women who are BB never become
totally bald like men do, but rather, their hair may become thin or sparse.
If two parents are heterozygous for baldness (note: that means he would be
bald, but she would not.), what are the chances of their children being
bald? Use a Punnett square to illustrate this.
Note: because the sex of a person does make a difference in how the
gene is expressed, you need to set this up as a dihybrid cross to account
for the sex of the children.
Baldness in humans is a dominant, sex-influenced trait. This gene is on the
autosomes, not the sex chromosomes, but how it is expressed is influenced
by the persons sex (due to hormones present, etc.). A man who is BB or Bb
will be bald and will be non-bald only if he is bb. A woman will only be
bald if she is BB and non-bald if she is Bb or bb (its almost like B is
dominant in males and b is dominant in females). Actually, because of the
influence of other sex-related factors, most women who are BB never become
totally bald like men do, but rather, their hair may become thin or sparse.
If two parents are heterozygous for baldness (note: that means he would be
bald, but she would not.), what are the chances of their children being
bald? Use a Punnett square to illustrate this.
Note: because the sex of a person does make a difference in how the
gene is expressed, you need to set this up as a dihybrid cross to account
for the sex of the children.

 In sweet peas, purple flower color (P) is dominant over white (p), but there
is also a control gene such that if the plant has a C, the purple has
permission to express itself. If the plant is cc, the purple does not
have permission to express itself and the flower will be white anyway. If
a plant with homozygous purple, controlled flowers is crossed with a plant
with white, non-controlled flowers, diagram the Punnett square for the
F1 and F2 generations and calculate the genotype and
phenotype ratios.
In sweet peas, purple flower color (P) is dominant over white (p), but there
is also a control gene such that if the plant has a C, the purple has
permission to express itself. If the plant is cc, the purple does not
have permission to express itself and the flower will be white anyway. If
a plant with homozygous purple, controlled flowers is crossed with a plant
with white, non-controlled flowers, diagram the Punnett square for the
F1 and F2 generations and calculate the genotype and
phenotype ratios. 